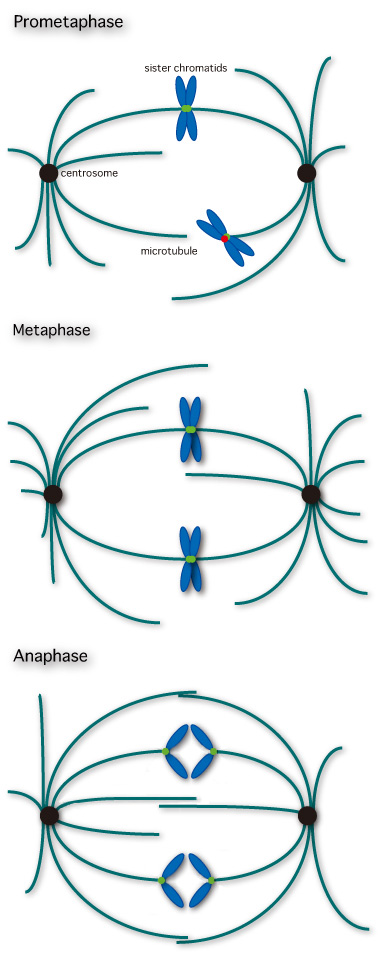
S期に複製された染色体は姉妹同士が対合して姉妹染色分体を形成します(図)。この対合は間期を通じて維持されます。有糸分裂初期から染色体有糸分裂期へ進行すると姉妹染色分体は凝縮を開始し、核の周辺では紡錘糸の形成極である中心体が二分割し、それぞれが核の両端へ移動をはじめます。核膜の崩壊に伴い紡錘糸と姉妹染色分体との活発な相互作用がおこります。姉妹染色分体の動原体にはそれぞれの極から伸長する紡錘糸が接続し、染色分体は紡錘糸に依存した運動により細胞の中心にある赤道面上に移動します。総ての姉妹染色分体が適切に紡錘糸と接続し赤道面上に整列すると、有糸分裂後期に移行し姉妹染色分体は解離します。有糸分裂前中期には、ある染色分体は両極から伸長した紡錘糸との接続、さらに赤道面上へ移動を完了している一方で、他の染色分体は紡錘糸との接続が完了していない状況がおこり得ます。このような状況では赤道面上にある姉妹染色分体は解離しません。すべての姉妹染色分体が赤道面に整列するまで染色分体の解離を抑制する機構、すなわちスピンドルチェックポイントが機能していると考えられています。
スピンドルチェックポイントはどのようにして姉妹染色分体の解離を抑制するのでしょうか?姉妹染色分体の対合はコヒーシンとよばれるタンパク質複合体により維持されています。有糸分裂後期にいたり染色分体解離する際にはコヒーシンの分断が必要です。セパレースは特異性の高いタンパク質分解酵素で、コヒーシンのサブユニットを分断します。セパレースの活性はその阻害因子、セキュリンにより制御されます。細胞周期のほとんどの時間で、セキュリンはセパレースに結合し、その活性を抑制します。有糸分裂後期になるとセキュリンはユビキチン化依存的に分解され、これにともないセパレースが活性化されます。すなわち、セキュリンの分解のタイミングが、姉妹染色分体の解離の時期を決定します。セキュリンのユビキチン化には、ユビキチンリガーゼであるAPC(Anaphase Promoting Complex)/Cyclosomeとその活性化に必要なCdc20(分裂酵母の相同体はSlp1)が必要です。スピンドルチェックポイントの機能因子の一つであるMad2はCdc20/Slp1と複合体を形成し、Cdc20/slp1によるAPCの活性化を阻害します。つまり、総ての動原体に紡錘糸が適切に結合するまで、スピンドルチェックポイントはCdc20/Slp1をターゲットとして、その活性を抑制することにより姉妹染色分体の解離を遅延させます(文献1,Kim et al., 1998)。
 |