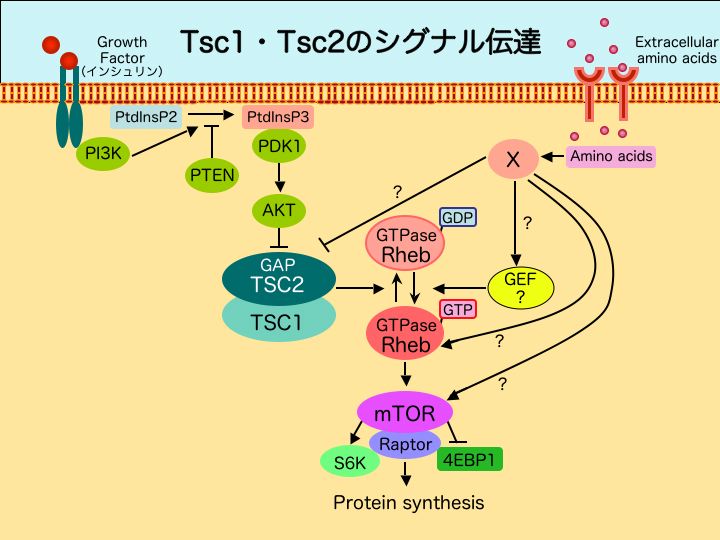
細胞を取りまく環境に栄養源が枯渇すると、細胞内ではいくつかのシグナル伝達系が活性化され、枯渇に適応するものと考えられています。
その一つは、Tsc1,Tsc2タンパク質によるものです。Tsc1タンパク質とTsc2蛋白質は複合体を形成し、インシュリン及びアミノ酸応答経路で機能することが知られています。Tsc2タンパク質はGAP(GTPase Activating Protein)として低分子量Gタンパク質(small GTPase)Rhebの活性を負に制御していると考えられています。Rheb GTPaseは、その下流因子であるmTORキナーゼを通してS6キナーゼや4EBP1を調節し、タンパク質合成を制御しています。すなわちTsc1/Tsc2を中枢とするシグナル伝達系は栄養状況や増殖因子の有無に呼応してグローバルなタンパク質合成量を制御し、細胞成長の調節をするものと考えられています(図参照)。
当研究室では、遺伝学的操作が容易な分裂酵母をモデル系として用い、Tsc1/Tsc2を中枢とするシグナル伝達系の全貌を明らかにすることを目的として研究を進めています。Tsc1またはTsc2を欠損する分裂酵母は、アミノ酸の取込み能力が顕著に低下します。また、通常は窒素源枯渇により誘導される減数分裂への進行が遅延します(文献3:Matsumoto, et al., 2002)。これらの表現型は、Tsc1,Tsc2タンパク質がグローバルなタンパク質合成量を制御するのみならず、アミノ酸取り込み酵素の活性や減数分裂誘導に必要な遺伝子発現をも制御していることを示唆します(文献4:Nakase et al., 2006)。
ヒトでTsc1,Tsc2に欠損がおこると結節性硬化症(Tuberous Sclerosis Complex;TSC)を発症します。TSCは多臓器(脳、目、心臓、腎臓、皮膚、肺)にわたり腫瘍形成をみる難治性疾患です。TSCの発症率は5,000~6,000人に1人であると報告されており、比較的頻度の高い遺伝性疾患です。我々の分裂酵母をモデル系とした研究が、将来の臨床応用に貢献することも多いに期待できます。